WikiDer > Митохондриальный противовирусный сигнальный белок






Митохондриальный противовирусный сигнальный белок (MAVS) это белок это важно для противовирусного врожденный иммунитет. MAVS расположен на внешней мембране митохондрий, пероксисом и эндоплазматического ретикулума (ER). При вирусной инфекции группа цитозольных белков обнаруживает присутствие вируса и связывается с MAVS, тем самым активируя MAVS. Активация MAVS заставляет инфицированную вирусом клетку секретировать цитокины. Это вызывает иммунная реакция который убивает инфицированные вирусом клетки хозяина, что приводит к удалению вируса.
Структура

MAVS также известен как стимулятор промотора I IFN-β (IPS-1), адаптер домена рекрутирования активации каспазы, индуцирующий I FN-β (CARDIF), или адаптер передачи сигналов, индуцированный вирусом (VISA).[5]. MAVS кодируется MAVS ген[5][6]. MAVS представляет собой белок из 540 аминокислот, который состоит из трех компонентов: N-концевого домена активации каспазы (CARD), богатого пролином домена и трансмембранного C-концевого домена (TM).[5].
После того, как ген MAVS был транскрибирован в РНК, рибосомы могут транслировать белок MAVS с двух разных сайтов.[5]. Начальный сайт трансляции генерирует полноразмерный белок MAVS. Альтернативный сайт трансляции генерирует более короткий белок, называемый «miniMAVS» или короткий MAVS (sMAVS).[5]. sMAVS - это белок MAVS из 398 аминокислот, в котором отсутствует домен CARD. Это важно, потому что в домене CARD два цитозольных белка связываются для активации MAVS, сигнализируя о том, что в клетке присутствует вирус.[5].
Функция
Вирусы с двухцепочечной РНК распознаются либо трансмембранным toll-подобным рецептором 3 (TLR3) или одним из двух цитозольных белков, геном I, индуцируемым ретиноевой кислотой (RIG-I) -подобные рецепторы и ген 5 (MDA5)[5][6][7][8]. RIG-I и MDA5 различаются вирусной РНК, которую они распознают, но они имеют много общих структурных особенностей, включая N-концевую карту CARD, которая позволяет им связываться с MAVS.[5]. Активация MAVS приводит к повышению уровня провоспалительных цитокинов за счет активации факторов транскрипции, ядерного фактора kB (NF-kB), регуляторный фактор интерферона 1 (IRF1) и регуляторный фактор интерферона 3 (IRF3)[5][6][7]. NFkB, IRF1 и IRF3 являются факторами транскрипции и играют критическую роль в продукции цитокинов.
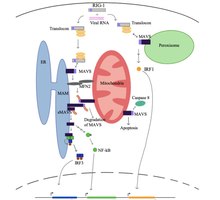
В состоянии покоя клетки белок, называемый митофузин 2 (MFN2), как известно, взаимодействует с MAVS, предотвращая связывание MAVS с цитозольными белками, такими как RIG-I и MDA5.[5][6]. Узнав вирус в цитозоле, митохондриально-ассоциированные мембраны ER (MAM) и митохондрии будут физически связаны с MFN2, а RIG-I связывается со вторым белком RIG-I с образованием белкового комплекса.[5][6][7]. Этот комплекс связывается с TRIM25 и молекулярный сопровождающий 14-3-3e, чтобы сформировать комплекс, называемый «транслокон»[5][6][7][8]. Транслокон перемещается в митохондрии, где он связывается с областью CARD на MAVS, что приводит к активации MAVS.[5][6][7][8]. Впоследствии белки MAVS связываются друг с другом через домен CARD и TM, чтобы задействовать несколько нижестоящих сигнальных факторов для формирования сигнального комплекса MAVS.[5][6]. Формированию этого сигнального комплекса MAVS помогают повышенные уровни митохондриальные активные формы кислорода (mROS), независимо от восприятия РНК[6][7]. Сигнальный комплекс MAVS взаимодействует с TANK-связывающая киназа 1 и / или протеинкиназы ИККА (ЧУК) и ИККБ (IKBKB), что приводит к фосфорилированию и ядерной транслокации IRF3[5]. Хотя трансдукция и регуляция сигнала MAVS до конца не изучены, активированные белки MAVS в митохондриях, ER и пероксисоме необходимы для максимального усиления противовирусного врожденного иммунного ответа.
Белок MAVS индуцирует апоптоз в инфицированных вирусом клетках-хозяевах путем взаимодействия с протеазой, называемой каспаза 8[5]. Активация апоптоза каспазой 8 не зависит от пути апоптоза Bax / Bak, основного пути апоптоза в клетках.[5].
Вирусное уклонение от MAVS
Некоторые вирусы, такие как цитомегаловирус человека (HCMV) и гепатит C (HCV), адаптировались для подавления функции MAVS в противовирусном врожденном иммунном ответе, способствуя репликации вируса.[5][9]. HCMV нарушает MAVS посредством локализованного в митохондриях вируса ингибитора белка апоптоза (vMIA), тем самым снижая провоспалительный цитокиновый ответ.[9]. vMIA также локализуется в пероксисоме, где vMIA взаимодействует с цитоплазматическим белком-шапероном Pex19, отключая транспортный механизм белков пероксисомальной мембраны.[9]. Штамм NS3-NS4A вируса гепатита C инактивирует передачу сигналов MAVS, расщепляя белок MAVS непосредственно перед направленным на мембрану доменом MAVS в MAM и пероксисоме, предотвращая передачу сигналов нижестоящего MAVS.[5].
Регулирование
Экспрессия и функция MAVS регулируются на транскрипционном, посттранскрипционном и посттрансляционном уровнях. На уровне транскрипции активные формы кислорода (АФК), образующиеся во время противовирусного ответа, действуют как отрицательный регулятор[5][6][7]. MAVS, кроме того, кодирует ряд вариантов сплайсинга, которые были предложены для регулирования MAVS. На посттранскрипционном уровне на MAVS присутствуют два сайта трансляции, которые могут генерировать два белка MAVS. Альтернативный сайт перевода находится выше по течению, что приводит к выражению sMAVS[5][6][7]. На трансляционном уровне белки, такие как семейство убиквитин E3 лигаза регулировать деятельность МАВС[5][6][7].
использованная литература
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000088888 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000037523 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v Васкес, Кристина; Хорнер, Стейси М. (15.07.2015). «Координация MAVS противовирусного врожденного иммунитета». Журнал вирусологии. 89 (14): 6974–6977. Дои:10.1128 / JVI.01918-14. ISSN 0022-538X. ЧВК 4473567. PMID 25948741.
- ^ а б c d е ж г час я j k л Моханти, Абхишек; Тивари-Пандей, Рашми; Панди, Нихар Р. (сентябрь 2019 г.). «Митохондрии: незаменимые участники врожденного иммунитета и хранители воспалительной реакции». Журнал сотовой связи и сигнализации. 13 (3): 303–318. Дои:10.1007 / s12079-019-00507-9. ISSN 1873-9601. ЧВК 6732146. PMID 30719617.
- ^ а б c d е ж г час я Jacobs, Jana L .; Койн, Кэролайн Б. (13 декабря 2013 г.). «Механизмы регуляции MAVS на митохондриальной мембране». Журнал молекулярной биологии. 425 (24): 5009–5019. Дои:10.1016 / j.jmb.2013.10.007. ISSN 0022-2836. ЧВК 4562275. PMID 24120683.
- ^ а б c Цзян, Цю-Син (2019). «Структурная изменчивость пути RLR-MAVS и чувствительное обнаружение вирусных РНК». Медицинская химия (Шарик (Объединенные Арабские Эмираты)). 15 (5): 443–458. Дои:10.2174/1573406415666181219101613. ISSN 1573-4064. ЧВК 6858087. PMID 30569868.
- ^ а б c Эшли, Кэролайн Л .; Абендрот, Эллисон; МакШарри, Брайан П .; Слободман, Барри (2019). «Интерферон-независимые врожденные ответы на цитомегаловирус». Границы иммунологии. 10: 2751. Дои:10.3389 / fimmu.2019.02751. ISSN 1664-3224. ЧВК 6917592. PMID 31921100.
дальнейшее чтение
- Нагасе Т., Исикава К., Кикуно Р., Хиросава М., Номура Н., Охара О. (2000). «Прогнозирование кодирующих последовательностей неидентифицированных генов человека. XV. Полные последовательности 100 новых клонов кДНК из мозга, которые кодируют большие белки in vitro». ДНК Res. 6 (5): 337–45. Дои:10.1093 / днарес / 6.5.337. PMID 10574462.
- Мацуда А., Сузуки Ю., Хонда Дж., Мурамацу С., Мацузаки О, Нагано И., Дои Т., Шимотохно К., Харада Т., Нисида Е., Хаяси Н., Сугано С. (2003). «Крупномасштабная идентификация и характеристика человеческих генов, активирующих сигнальные пути NF-kappaB и MAPK». Онкоген. 22 (21): 3307–18. Дои:10.1038 / sj.onc.1206406. PMID 12761501.
- Каваи Т., Такахаши К., Сато С., Кобан С., Кумар Х., Като Х., Исии К.Дж., Такеучи О, Акира С. (2005). «IPS-1, адаптер, запускающий RIG-I- и Mda5-опосредованную индукцию интерферона I типа». Nat. Иммунол. 6 (10): 981–8. Дои:10.1038 / ni1243. PMID 16127453. S2CID 31479259.
- Мейлан Э., Курран Дж., Хофманн К., Морадпур Д., Биндер М., Бартеншлагер Р., Чопп Дж. (2005). «Кардиф - это адаптерный белок в антивирусном пути RIG-I, на который нацелен вирус гепатита С». Природа. 437 (7062): 1167–72. Дои:10.1038 / природа04193. PMID 16177806. S2CID 4391603.
- Ли XD, Сун Л., Сет РБ, Пинеда Дж., Чен З.Дж. (2006). «Протеаза вируса гепатита С NS3 / 4A отщепляет митохондриальный противовирусный сигнальный белок от митохондрий, чтобы избежать врожденного иммунитета». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 102 (49): 17717–22. Дои:10.1073 / pnas.0508531102. ЧВК 1308909. PMID 16301520.
- О ДжХ, Ян Джо, Хан И, Ким М.Р., Бён СС, Чон Й.Дж., Ким Дж.М., Сон К.С., Но С.М., Ким С., Ю HS, Ким Ю.С., Ким Н.С. (2006). «Транскриптомный анализ рака желудка человека». Мамм. Геном. 16 (12): 942–54. Дои:10.1007 / s00335-005-0075-2. PMID 16341674. S2CID 69278.
- Лоо Ю.М., Оуэн Д.М., Ли К., Эриксон А.К., Джонсон К.Л., Фиш ПМ, Карни Д.С., Ван Т., Исида Х., Йонеяма М., Фудзита Т., Сайто Т., Ли В.М., Хагедорн С.Х., Лау Д.Т., Вайнман С.А., Лимон С.М. , Гейл М. (2006). «Вирусный и терапевтический контроль стимулятора 1 промотора IFN-бета во время инфицирования вирусом гепатита С». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 103 (15): 6001–6. Дои:10.1073 / pnas.0601523103. ЧВК 1458687. PMID 16585524.
- Ченг Дж., Чжун Дж., Чисари Ф.В. (2006). «Ингибирование дцРНК-индуцированной передачи сигналов в клетках, инфицированных вирусом гепатита С, с помощью NS3-протеазозависимых и независимых механизмов». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 103 (22): 8499–504. Дои:10.1073 / pnas.0602957103. ЧВК 1482521. PMID 16707574.
- Лин Р., Лакост Дж., Нахаи П., Сан К., Ян Л., Паз С., Уилкинсон П., Юлкунен И., Витур Д., Мерс Э., Хискотт Дж. (2006). «Диссоциация молекулярного комплекса MAVS / IPS-1 / VISA / Cardif-IKKepsilon от внешней мембраны митохондрий протеолитическим расщеплением NS3-4A вируса гепатита С». Дж. Вирол. 80 (12): 6072–83. Дои:10.1128 / JVI.02495-05. ЧВК 1472616. PMID 16731946.
- Саха С.К., Пьетрас Э.М., Хе Дж.К., Кан-младший, Лю С.Ю., Оганесян Г., Шахангиан А., Зарнегар Б., Шиба Т.Л., Ван Й., Ченг Дж. (2006). «Регулирование противовирусных реакций за счет прямого и специфического взаимодействия между TRAF3 и Cardif». EMBO J. 25 (14): 3257–63. Дои:10.1038 / sj.emboj.7601220. ЧВК 1523175. PMID 16858409.
- Босолей С.А., Виллен Дж., Гербер С.А., Раш Дж., Гиги С.П. (2006). «Вероятностный подход к высокопроизводительному анализу фосфорилирования белков и локализации сайтов». Nat. Биотехнология. 24 (10): 1285–92. Дои:10.1038 / nbt1240. PMID 16964243. S2CID 14294292.
- Опиц Б., Винцинг М., ван Лаак В., Шмек Б., Хайне Г., Гюнтер С., Прейсснер Р., Слефогт Х., Н'Гессан П. Д., Эйтель Дж., Гольдман Т., Флигер А., Сутторп Н., Гиппенштиль С. (2007). «Legionella pneumophila индуцирует IFNbeta в эпителиальных клетках легких через IPS-1 и IRF3, которые также контролируют репликацию бактерий». J. Biol. Chem. 281 (47): 36173–9. Дои:10.1074 / jbc.M604638200. PMID 16984921.
- Чен З, Бенюро Й, Райнбранд Р., Йи Дж, Ван Т., Уортер Л., Ланфорд Р. Э., Вайнман С. А., Лимон С. М., Мартин А., Ли К. (2007). «GB вирус B нарушает передачу сигналов RIG-I посредством NS3 / 4A-опосредованного расщепления адапторного белка MAVS». Дж. Вирол. 81 (2): 964–76. Дои:10.1128 / JVI.02076-06. ЧВК 1797450. PMID 17093192.
- Хирата Y, Броке AH, Menchén L, Kagnoff MF (2007). «Активация защитных механизмов врожденного иммунитета посредством передачи сигналов через RIG-I / IPS-1 в эпителиальных клетках кишечника». J. Immunol. 179 (8): 5425–32. Дои:10.4049 / jimmunol.179.8.5425. PMID 17911629.
- Цзэн В., Сюй М., Лю С., Сунь Л., Чен З.Дж. (2009). «Ключевая роль полиубиквитинирования Ubc5 и лизина-63 в вирусной активации IRF3». Mol Cell. 36 (3): 315–25. Дои:10.1016 / j.molcel.2009.09.037. ЧВК 2779157. PMID 19854139.
- Лю С., Чен Дж, Цай Х, Ву Дж, Чен Икс, Ву И, Сун Л, Чен ЗДж (2013). «MAVS привлекает несколько лигаз убиквитина E3 для активации противовирусных сигнальных каскадов». eLife. 2 (e00785): e00785. Дои:10.7554 / eLife.00785. ЧВК 3743401. PMID 23951545.
- Лю С., Цай Х, Ву Дж, Конг Q, Чен Х, Ли Т, Ду Ф, Рен Дж, Ву И, Гришин Н., Чен З. Дж. (2015). «Фосфорилирование адаптерных белков врожденного иммунитета MAVS, STING и TRIF вызывает активацию IRF3». Наука. 347 (6227): ааа2630. Дои:10.1126 / science.aaa2630. PMID 25636800.
Эта статья включает текст из Национальная медицинская библиотека США, который находится в всеобщее достояние.